Модели и реальность
Конечно, такая слабая эскизная модель, как (44), не может использоваться для конкретных расчетов, она предназначена не для этого, а только для организации мышления и упорядоченного обсуждения участия граничных поверхностей в формировании структуры океана. Поэтому она не может быть признана удовлетворительной при рассмотрении объекта с позиций частных океанологических наук: биологии, химии и особенно физики океана, поскольку основанная на максимальной генерализации, она преднамеренно игнорирует важные конкретно-специфические черты, составляющие интерес соответствующих частных наук. Эта модель никак не передает богатства оттенков явления, но может внести ясность при условии, если она обладает общностью и позволяет описать в самых грубых чертах распределение разнообразных типовых конкретных характеристик.
Первое, что можно отметить, это некоторое общее совпадение, как видно из модельных описаний, с вертикальным разделением океана на гидрологические структурные зоны [Степанов, 1974]. Что касается распределения компонент, то на рис. 14 оно неплохо отражается моделью для случаев увеличения субстанции с глубиной (соленость, плотность, продукты минерализации) и для случая уменьшения субстанции с глубиной (скорости механических, химических, биохимических процессов, концентрации органических веществ, биомассы).
Легко заметить, что физические, гидромеханические, химические и особенно биологические характеристики активности изменяются в верхнем пограничном слое по сравнению с инертным неодинаково. Различия, по-видимому, возрастают с усложнением формы движения, т. е. намечается связь с иерархией уровней и соподчинением форм движения от механической к биологической, из которых первая форма относительно независима от остальных, а высшие зависят от низших. Поэтому биологические характеристики могут служить хорошим индикатором граничных поверхностей.
Горизонтальные профили распределения биомассы от таких разных границ, как берега континентов (рис. 15) и островов, экватор, кромка льдов, фронты, показывают сходную картину и не сильно отличаются от того, что дает примитивная эскизная модель (44).
![Рис. 14. Сравнение модели и типового распределения относительных характеристик. 1 - среднее значение для океана, по 'Физике океана', 1978; 2 - 3 - по 'Океанографической энциклопедии', 1974; 4 - по Р. Стренбергу, 1974; 5 - по Б. А. Скопенцева, 1975; 6, 9 - Атлантический полигон-70; 7, 8 - по Ю. Ф. Лукашова, 1973; 10 - по Г. Дрью [Л. А. Зенкевич, 1951]; 11 - по Е. Генштелю, 1938. За единицу взяты значения характеристик в удаленной от границ области, Г - граница разных водных масс.](pic/000089.jpg)
Рис. 14. Сравнение модели и типового распределения относительных характеристик. 1 - среднее значение для океана, по 'Физике океана', 1978; 2 - 3 - по 'Океанографической энциклопедии', 1974; 4 - по Р. Стренбергу, 1974; 5 - по Б. А. Скопенцева, 1975; 6, 9 - Атлантический полигон-70; 7, 8 - по Ю. Ф. Лукашова, 1973; 10 - по Г. Дрью [Л. А. Зенкевич, 1951]; 11 - по Е. Генштелю, 1938. За единицу взяты значения характеристик в удаленной от границ области, Г - граница разных водных масс.
Сгущение жизни в пограничных слоях даже качественно не всегда удается объяснить влиянием температуры, освещенности, биогенных элементов, и еще реже влияние этих факторов позволяет дать количественное объяснение. Если докапываться до истины, то на отдельных границах раздела обнаруживаются самые разнообразные конкретные фундаментальные свойства и явления, которые в конечном счете влияют на формирование конкретных экологических систем. Это обращение в нуль коэффициента турбулентной диффузии на твердой и свободной поверхностях или параметра Корио-лиса на экваторе; поглощение солнечной радиации в верхнем пограничном слое и «выедание» в нем фитопланктоном биогенных элементов; вертикальные токи в районах вергенций и ап-веллингов и ускорение биогеохимических циклов в последних; поступление питательных веществ и отвод метаболитов; размножение и гибель организмов; адсорбция, метаморфиза-ция состава, обмен многочисленными компонентами, полный список которых неизвестен, и многое другое.
Но, может быть, за всеми этими свойствами граничных поверхностей и явлениями в них и стоит именно разнообразие условий существования живых организмов как общий признак возможности сгущения жизни и активизации экологических систем? Условий, достаточно разнообразных, чтобы активно размножаться, питаться и даже активно погибать, ведь и гибель - важное звено экологических и биогеохимических циклов. Может быть, к ускорению этих циклов и придется в конце концов свести биологический эффект граничных поверхностей?
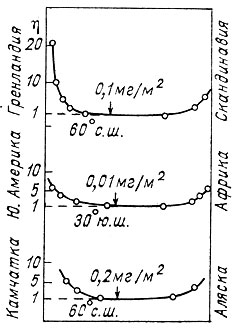
Рис. 15. Величина относительной биомассы зоопланктона на разных широтах океана. За единицу взято значение биомассы на большом удалении от берега.
Можно наметить схему, по которой об этом удобно говорить математическим языком, добиваясь общности описания. Скорее всего, это должны быть языки теории множеств, математической логики, теории вероятности. Обозначим лишь путь к такому разговору.
Мы уже характеризовали множеством

водную массу М. Пусть и окружающие ее объекты Ej (атмосфера, дно, берег, инородные воды и т. д.) характеризуются неэквивалентными между собой множествами
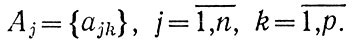
Элементы множеств
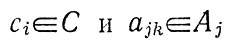
- физические, химические и биологические параметры.
По обе стороны границы раздела
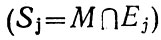
множества имеют общие элементы, т. е. их пересечение не составляет пустое множество
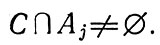
Однако только пограничные слои характеризуются объединением (суммированием U) множеств
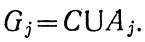
Поэтому разнообразия условий, а если с ними связывать активность экологических систем, то и вероятность сгущения жизни Р, резко возрастают при переходе от батипелагиали (удаленных от всех внешних границ раздела глубоководных слоев) к пограничным слоям пела-гиали, примыкающим к одинарным границам раздела океан - атмосфера или вода - дно. Возрастание продолжается к пограничным слоям, примыкающим к многократным границам раздела (пересечению),
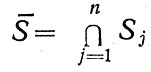
которые характеризуются объединением множеств
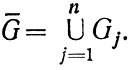
Так, интегральные характеристики активности экологических систем - время оборота органического вещества и биохимическое потребление кислорода, составляющие в глубинах океана величины порядка 103 лет и 10-3 мл О2/л в год соответственно [Скопинцев, 1975],- изменяются на 3-4 порядка при переходе к верхнему пограничному слою в открытом море и еще на 1-2 порядка в прибрежных районах. При этом удельная активность (активность одного организма) может даже падать, т. е. речь идет об активности именно экологических систем, а не организмов.
Таким образом, начало формализованного разговора получается. Для того чтобы его продолжить, надо найти способ формализованного описания разнообразия условий.
Для начала М и Ej расмотрим как взаимодействующие элементы, образующие локальную систему. Число состояний системы N характеризуем самым простым способом (тоже на первых порах): наличием или отсутствием каждой из двух возможных связей между двумя элементами (прямой и обратной). С ростом числа элементов n+1(n объектов плюс вода) число возможных состояний сильно возрастает:
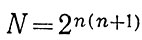
Если в качестве меры разнообразия условий R принять простейшую, например типа меры Р. Хартли
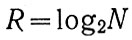
то для грубой оценки примем выражение
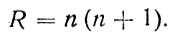
(46)
Трудно найти вид функции P(R), где Р-вероятность сгущения жизни. Для сравнения двух районов можно использовать приблизительное соотношение
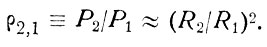
(47)
В соотношении (47) квадрат, отражает наличие положительной обратной связи в цепочке поверхности раздела - живое вещество.
На рис. 16 даны наиболее существенные внешние и внутренние границы раздела, а также ориентировочные оценки разнообразия условий R и относительной вероятности сгущения жизни р. Хорошо видны изменения разнообразия условий в районе банки и по мере приближения к шельфу. Вблизи шельфа условия максимально разнообразны, наиболее вероятно сгущение жизни, максимальна активность всех процессов.
Берег можно назвать многократной границей, на которой пересекаются или сближаются поверхности раздела (они же поверхности обмена) разной природы: вода-воздух, вода - суша, вода - дно, вода - взвешенное вещество, вода - живое вещество и др. В свою очередь в активной береговой зоне, где существуют и другие каналы обмена - устья рек, каньоны, апвеллинги, проливы,- возникают точки и очаги повышенной активности. Близость сероводородной зоны, судя по модели, также повышает вероятность сгущения жизни в рядом расположенных кислородных зонах, несмотря на то, что сама она почти безжизненна. Что это - издержки формализации? По-видимому, нет. Известен, по крайней мере, один механизм, вызывающий такое явление: в сероводородной зоне происходит мобилизация фосфатов из донных отложений. Модель циркумграничной структуры океана подводит к еще одной проблеме - проблеме биогеохимического фронта.
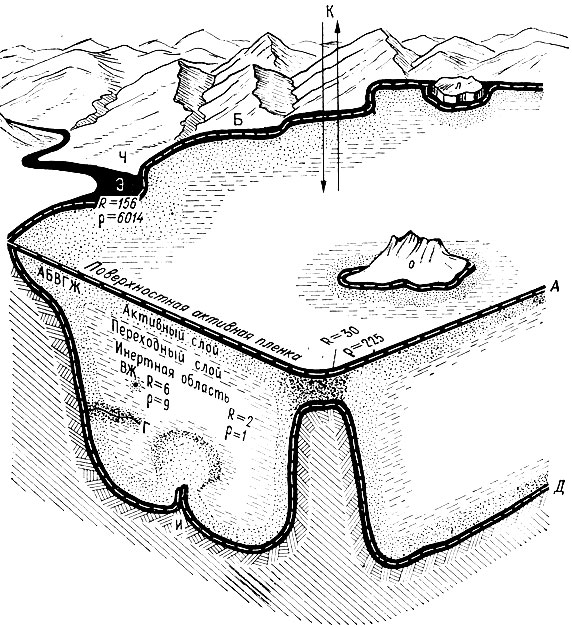
Рис. 16. Циркумграничная структура океана. Дана оценка разнообразия условий R и относительной вероятности сгущения жизни р. Масштаб сильно искажен. Наиболее существенные внешние и внутренние границы раздела: А - с атмосферой, К - космосом, Б - берегом, В - взвешенным веществом, Г - иной водной массой, Д - дном, Ж - живым веществом, И - глубинным веществом дна (источники, вулканы), Л - льдом, О - островом, Ч - антропогенным веществом (человек - океан), Э - речными водами (эстуарии).
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://underwater.su/ 'Человек и подводный мир'