Глава 4. Технология разработки и воспроизводства морских биологических ресурсов
4.1. Количественная оценка запасов промысловых организмов
"Океаны и моря,- писал академик Л. А. Зенкевич,- представляют собой в наиболее ярко выраженной степени целостный природный объект с тесным взаимодействием и взаимообусловленностью всех протекающих в нем процессов". Вся совокупность населяющих моря и океаны живых организмов образует биологические ресурсы Мирового океана. В результате роста и развития этих организмов происходит новообразование биомассы - биологическое продуцирование, об уровне которого судят по величине продукционных показателей. Основной из них - масса живого вещества, воспроизведенного в водоеме за единицу времени.
Биологическая продуктивность Мирового океана в значительной степени зависит от океанологических характеристик его районов, рельефа дна, баланса биогенных элементов и особенностей продуцирования фито- и зоопланктона [37]. Поэтому для получения полной ее характеристики необходимы длительные комплексные исследования. Значительно легче оценить суммарную промысловую продукцию, которая на один - три порядка ниже общей продукции беспозвоночных, планктона и бентоса.
Другой важнейший показатель биологических ресурсов океана - запас, т. е. совокупность всех видов организмов, которые являются или могут стать предметом промысла. В последние годы все большее внимание уделяется определению запасов непромысловых и хозяйственно малоценных объектов: кормовых (для рыб или промысловых беспозвоночных), вредных - врагов или конкурентов промысловых объектов, организмов-мелиораторов и др.
Научная организация использования биологических ресурсов невозможна без конкретной оценки запаса и учета количественных изменений, происходящих с ним под влиянием промысла. Рациональный промысел базируется на следующих принципах: структура промыслового стада есть результат взаимодействия пополнения, роста и убыли трансформируемых через среду обитания животных; необходимое соответствие между запасом и выловом (оптимальный улов) можно получить только путем селективного промысла, специфического для каждого вида и условий его существования.
Промысловое стадо изучается путем отбора проб. Для их анализа широко используются методы математической статистики и теории выборочных наблюдений, позволяющие получить сведения о генеральной совокупности. Наряду с этими методами получили распространение методы построения математических моделей, характеризующих состояние промыслового стада. Однако необходимо отметить, что теория использования промысловых запасов разработана в основном для рыб; многие важные вопросы, присущие хозяйственному использованию морских беспозвоночных и водорослей, еще ждут решения.
К настоящему времени получена общая картина количественного распределения в Мировом океане первичной продукции и зоопланктона, населяющего поверхностный 100-метровый слой океанических вод. Сведения о количественном распределении других пелагических организмов (фитопланктон, нектон, население больших океанских глубин) все еще ограничены.
Количественные исследования населения морей и океанов стали возможными только после появления орудий количественного сбора - планктонной сетки, изобретенной в 1877 г. немецким планктонологом Гензеном, и дночерпателей, введенных в практику морских исследований в 1910 г. датским зоологом Петерсоном. И в настоящее время дночерпатель служит основным орудием количественных сборов бентоса: на дночерпательных пробах, полученных с площади 0,2-0,25 м2, основано большинство известных данных о биомассе бентоса. Однако количественные методы сбора донной фауны дночерпателями весьма несовершенны, и полученные с их помощью показатели значительно занижены по сравнению с фактическими. Вследствие небольшой площади дночерпателя в него обычно не попадают крупные животные, распределенные на дне разреженно и неравномерно, а также быстро двигающиеся животные, спугиваемые движением воды перед падающим дночерпателем. Более точные данные о численности организмов, обитающих на поверхности грунта, дает траловая съемка, но она не позволяет учитывать закапывающихся животных, численность и биомасса которых довольно значительны. Наиболее достоверный количественный учет донных организмов в верхней сублиторали может быть выполнен с помощью водолазов. На мягких грунтах биомасса, определенная водолазами, оказывается в 1,5-3 раза больше по сравнению с данными дночерпательных проб. На твердых грунтах (камни, скалы) эти показатели могут различаться на порядок и больше.
Существующие в настоящее время методы определения запаса можно разделить на две группы: методы прямого подсчета и методы оценки относительных показателей, косвенно характеризующих состояние запаса.
В основе прямых методов лежит предположение о том, что промысловые объекты в течение заданного промежутка времени равномерно распределены по рассматриваемой промысловой площади. Получаемая информация основывается на визуальном методе учета, использовании количественных орудий лова, применении фото-, кино- и телеустановок, подсчете численности отложенных икринок, гидролокации, использовании результатов мечения, определения величины уловов. Косвенные методы в отличие от прямых не позволяют вычислить промысловый запас в массовом или штучном выражении, но дают возможность судить о характере изменений численности и состава промыслового стада по изменению тех или иных показателей его биологического процветания или угнетения. Очевидно, что любые методы должны учитывать особенности биологии данного вида, структуру ареала, связь между ареалом и промысловыми полями.
Один из наиболее широко используемых прямых методов оценки запаса - определение улова при разведочном тралении (траловая съемка). В основе обработки результатов такой съемки лежит принцип случайной выборки. Численность популяции определяется по формуле Грослайна
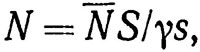
(4.1)
где 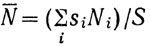 - осредненный улов с единицы площади (индекс численности); S - общая площадь обследуемой акватории; γ - абсолютный коэффициент уловистости; s - протраленная площадь. Чтобы повысить представительность получаемых данных, необходимо при расчете учесть дисперсию индекса численности. Советскими исследователями* предложено оценивать эту величину как сумму вида
- осредненный улов с единицы площади (индекс численности); S - общая площадь обследуемой акватории; γ - абсолютный коэффициент уловистости; s - протраленная площадь. Чтобы повысить представительность получаемых данных, необходимо при расчете учесть дисперсию индекса численности. Советскими исследователями* предложено оценивать эту величину как сумму вида
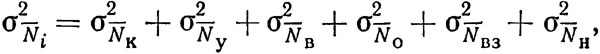
(4.2)
где отдельные слагаемые - вариации, обусловленные колебаниями плотности распределения промысловых объектов 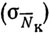 , изменением уловистости
, изменением уловистости 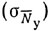 , временем суток
, временем суток 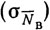 , опытом экипажа
, опытом экипажа 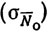 , взаимодействием перечисленных факторов
, взаимодействием перечисленных факторов 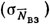 и неучтенными факторами
и неучтенными факторами 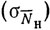 .
.
* (Биденко Г. Е. и др.- Тр. АтлантНИРО, Калининград, 1977, вып. 73, с 30-38.)
Для оценки плотности скоплений пелагических рыб успешно используются акустические методы. Разреженные скопления изучаются с помощью высокочастотных эхолотов, обладающих хорошей разрешающей способностью. Для изучения плотных скоплений применяют низкочастотные эхолоты. Часто оценка плотных скоплений требует комплексного использования акустических и фотографических методов.
Для учета пелагических и особенно донных скоплений рыб и беспозвоночных широко применяется притраловая фотограмметрическая съемка - особый вид измерительной фотосъемки, обеспечивающей точную дешифровку снимков. При такой съемке приходится учитывать влияние самого трала на результаты учета. Для определения этого влияния проводились сравнительные визуальные оценки плотности невозмущенного скопления рыб и скопления на входе в трал [18]. Было установлено, что в зоне съемки плотность скопления увеличивается. Это объясняется двумя причинами: предварительной концентрацией рыбы под воздействием турбулентности, создаваемой элементами, движущимися перед тралом, и движением рыбы внутри трала в направлении траления. Данные сравнительной оценки позволяют определить коэффициенты концентрации объектов для различных типов тралов и внести соответствующие поправки в расчеты.
Чтобы вести прямое фотографическое измерение плотности скоплений объектов лова без учета поправок на указанные возмущения, необходимо выносить регистрирующую аппаратуру за пределы зоны сильных возмущений. Для этого рекомендуется использовать специальный трос (фальшподбору), натянутый между траловыми досками, обеспечивающими горизонтальное раскрытие трала (вертикальное раскрытие создается с помощью поплавков и грузов). На центральной части фальшподборы устанавливают две-три фотокамеры типа "Тритон" или "Тритон-2". Благодаря малым размерам и импульсному характеру подсветки буксируемые на трале фотокамеры меньше пугают рыб, чем обитаемые аппараты или телевизионные устройства с постоянным освещением [52].
Если же необходимо исследовать поведение объекта и выработать методику определения плотности его распределения, то здесь незаменимы обитаемые подводные аппараты, а на малых глубинах - водолазы. Визуальные наблюдения позволяют с наибольшей точностью изучать распределение крупных донных рыб и беспозвоночных, ведущих одиночный образ жизни и не реагирующих на присутствие подводного аппарата или водолаза.
Особенности визуальной оценки плотности скоплений рассмотрены в работе [18]. Автор описывает методику применительно к наблюдениям донных скоплений с помощью подводного аппарата, однако полученные закономерности могут быть применены и к наблюдениям, выполненным водолазами. При движении наблюдателя вблизи дна просмотренная им площадь
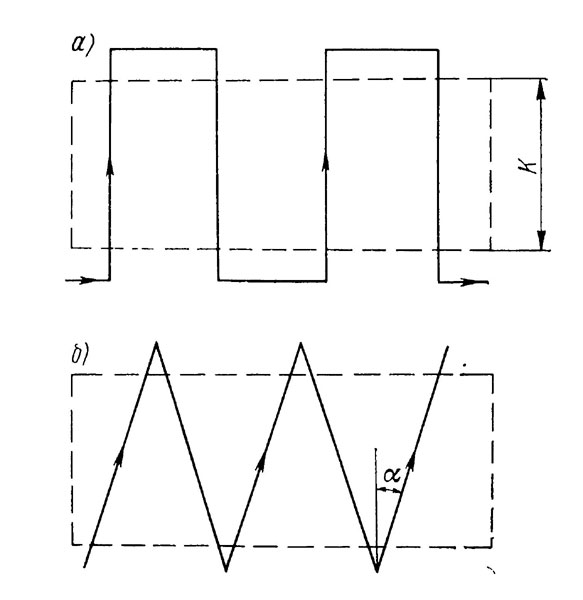
(4.3)
где h - расстояние от иллюминатора подводного аппарата до дна, υ - скорость движения, t - время наблюдения, β - половинный угол зрения из иллюминатора, α - угол отклонения направления визирования от вертикали.
Просматриваемый объем воды в таком случае
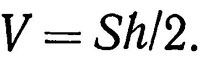
(4.4)
При движении подводного аппарата в толще воды вдали от дна просмотренный объем воды
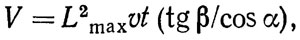
(4.5)
где Lmax - дальность видимости в воде.
Расстояние h определяется с помощью эхолота, установленного на подводном аппарате, оптическим дальномером или визуально. Значения углов находят расчетным путем. Дальность видимости Lmax, как правило, определяется визуально, для чего необходима специальная тренировка наблюдателя на тест-объектах. По окончании наблюдения определяют среднюю плотность скопления каждого вида: для донного распределения δs = N/S, для объемного распределения δv= N/V.
При оценке достоверности результатов необходимо учитывать особенности поведения подвижных организмов по отношению к наблюдателю. Так, ориентирование рыб в случайных направлениях может свидетельствовать об отсутствии влияния подводного аппарата на их поведение.
Проводя учет рыб из подводного аппарата, а затем траление с притраловой фотосъемкой по тому же маршруту (или чередуя спуски и траления на одном скоплении), можно подсчитать коэффициент концентрирования рыб тралом и коэффициенты уловистости. Эти показатели в дальнейшем используются для определения абсолютных плотностей скоплений по фотосъемкам или уловам без дополнительных спусков подводных аппаратов.
Рассмотрим вопрос о величине необходимой выборки, имеющий важнейшее значение при учете донных организмов независимо от используемого метода. Будем считать, что учитываемые организмы распределены на грунте случайным образом; это вполне допустимо при учете крупных и достаточно редко распределенных животных, к которым относится большинство промысловых организмов. Плотность распределения объектов численностью N на некоторой площади S определяется формулой δ = N/S. Дисперсия величины δ складывается из дисперсий определения средних значений случайных величин N и S. Поэтому для доверительных интервалов можно записать
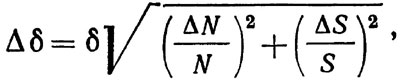
(4.6)
где Δδ, ΔN, ΔS - доверительные интервалы соответственно для δ, N и S.
Для относительных погрешностей εδ=Δδ/δ, εN = ΔN/N и εS= ΔS/S получим
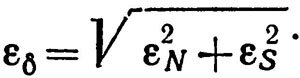
(4.7)
Количество экземпляров, попадающее в случайно выбранную область, распределено по закону Пуассона. Пусть на дно m раз брошена учетная рамка, а количество попавших в нее животных оказалось равным х1, х2, х3, ..., xm. При пуассоновском распределении нет необходимости знать отдельные значения χ, достаточно определить сумму 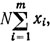 , т. е. общее количество подсчитанных особей. На основании этого можно построить график (рис. 4.1), определяющий доверительные интервалы для среднего значения плотности распределения объектов при различной относительной погрешности (величина εδ определена по литературным данным).
, т. е. общее количество подсчитанных особей. На основании этого можно построить график (рис. 4.1), определяющий доверительные интервалы для среднего значения плотности распределения объектов при различной относительной погрешности (величина εδ определена по литературным данным).
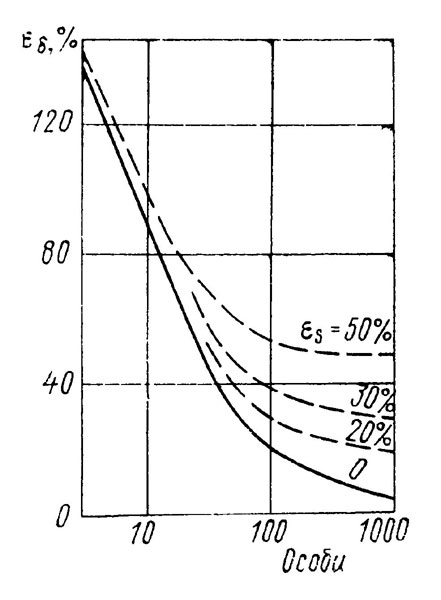
Рис. 4.1 Погрешность определения плотности распределения организмов в зависимости от величины выборки
Для оценки запасов организмов, обитающих на мелководье, широко используют водолазов. В этом случае животные подсчитывайте или на точно определяемой площади, обычно внутри укладываемой на дно рамки заданного размера, или на приближенно определяемой площади, например, на полосе, направление которой задается протянутым по дну фалом, или даже на неучтенной площади, когда фиксируется только время движения пловца.
Для оценки границ применимости разных методов обратимся к рис. 4.1. Видно, что при малых N (менее 20) значение εδ определяется главным образом величиной выборки и мало зависит от εS. Отсюда следует, что применение пробных рамок предпочтительнее при учете животных с большой плотностью распределения (выше 0,2 особи/м2). При подсчете организмов с малой плотностью распределения преимущество имеют методы, позволяющие увеличить объем выборки путем быстрого просмотра большой площади и существенно повысить эффективность учета. Так, при учете методом многократных галсов пловец движется прямыми галсами на определенном расстоянии от дна. Подсчет животных проводится на полосе, ширина которой определяется полем зрения маски или нанесенными на стекло двумя визирными линиями. Длину пройденного пути подсчитывают с помощью лага, по времени движения или по числу выполненных галсов. Расстояние от дна и средняя скорость движения для опытного водолаза весьма стабильны, и ошибка в их определении не превышает 10 %.
Если форма участка учета близка к кругу, то пловец движется произвольными галсами, меняя курс на границе участка. На лишенном ориентиров дне или в малопрозрачной воде точка поворота определяется натяжением сигнального конца, которым пловец связан с лодкой; при работе в очень прозрачной воде с заметными ориентирами граница участка легко определяется визуально.
Просмотренная площадь составит
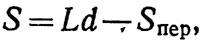
(4.8)
где Sпер - площадь всех пересечений галсов (участков, проходимых дважды), L - длина пути, d - ширина учетной полосы.
Зависимость средней относительной площади пересечений S'=Sпep/d2 от относительной длины пути L/R (участок учета - круг радиусом R) можно представить графически (рис. 4.2). Истинное количество особей на просмотренной площади определяется разностью Nист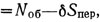 , или
, или 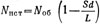 , где Nоб - подсчитанное пловцом число особей. Учет на участке заканчивается, когда истинное количество особей, вычисленное по этой формуле, обеспечит желаемую точность (см. рис. 4.1). При большом объеме исследований и достаточном опыте отрабатываются наиболее удобные параметры учета d, L и R, что позволяет использовать упрощенное выражение
, где Nоб - подсчитанное пловцом число особей. Учет на участке заканчивается, когда истинное количество особей, вычисленное по этой формуле, обеспечит желаемую точность (см. рис. 4.1). При большом объеме исследований и достаточном опыте отрабатываются наиболее удобные параметры учета d, L и R, что позволяет использовать упрощенное выражение 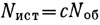 , где с - постоянная для данных условий величина.
, где с - постоянная для данных условий величина.
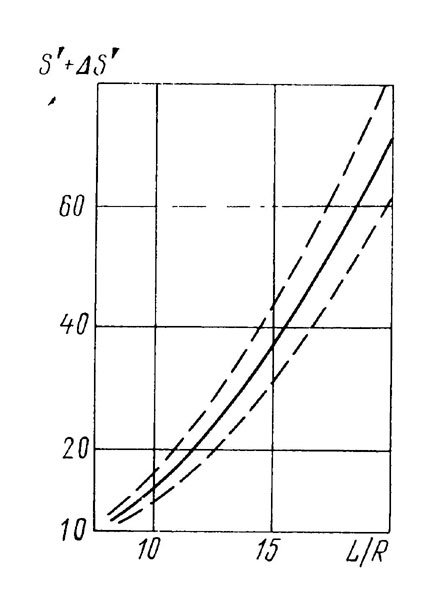
Рис. 4.2. Зависимость относительной площади пересечений (S') от относительной длины пути (L/R) на участке учета, имеющем форму круга
Если животные подсчитываются на участке в виде полосы, то пловец движется зигзагообразными галсами. При этом точки поворота должны быть расположены за пределами скопления организмов (рис. 4.3). Если на участке шириной k сделано q галсов, то для средней плотности распределения получим δ = N/qdk (рис. 4.3, а) и δ= N cosα/qdk (рис. 4.3,6).
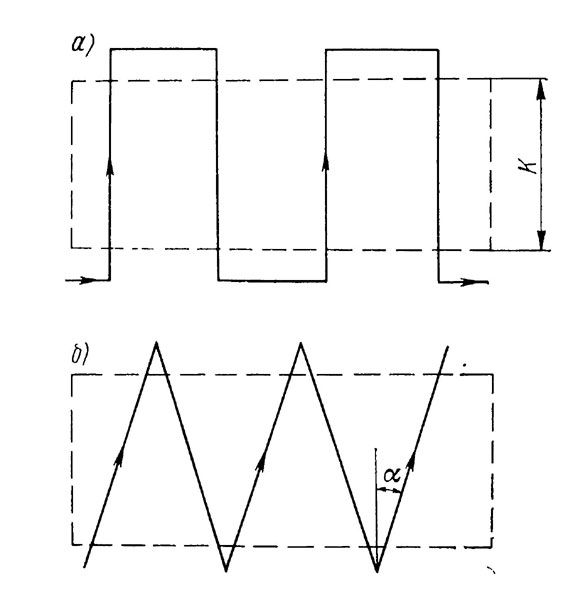
Рис. 4.3. Учет на площадке в форме вытянутой полосы
Во многих случаях запас наиболее удобно оценивать по результатам сбора. Для определения плотности распределения организмов с помощью этого метода следует воспользоваться формулами, приведенными в § 4.3.
Для учета запасов морских растений и донных животных, образующих однородные скопления, можно использовать метод линейного пересечения Кенфилда, широко применяющийся в геоботанике. Здесь в качестве учетных единиц используются случайно расположенные линейные трансекты равной длины, на которых измеряются отрезки их пересечения с пятнами изучаемого объекта. Учет ведется по формуле
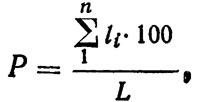
(4.9)
где Р - доля занятой площади, %; li - отрезки пересечения; L - длина трансекты.
Длина и количество используемых трансект зависит от рельефа дна и характера распределения учитываемого объекта [15].
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://underwater.su/ 'Человек и подводный мир'