Глава III. Чтобы плавать быстрее рыб...
Китообразные относятся к наиболее быстроходным обитателям гидросферы и превосходят в скорости рыб, которыми они питаются. Рекордсменом-скоростником в подотряде усатых китов считался полосатик сейвал, который в рывках при легком ранении будто бы мог достигать скорости 65 км/ч. Но это основывалось лишь на приблизительной оценке подвижности животных во время китобойного промысла, где точных замеров никто не проводил. Известно, что турбулентность, или завихрения, в пограничном слое жидкости, задерживающая движение, увеличивается не только со скоростью, но и с размером движущегося тела. Поэтому мелкие китообразные в особых случаях могут показывать резко преувеличенную (ложную) скорость хода. Например, дельфины, оседлывая корабельные волны и используя толкающую гидродинамическую силу, пристраивались к носу быстроходных эсминцев, двигавшихся со скоростью 65 км/ч. Но это неестественная скорость дельфинов.
Гораздо более надежные данные получены опытным путем. В 1966 г. гидродинамики США Томас Ланг и Карин Прайор провели эксперименты в лагуне на Гавайских островах над малайскими продельфинами, обученными гоняться за плавучей приманкой, которую быстро перемещали по поверхности воды с помощью электрической лебедки. Так установили максимальную скорость для этих дельфинов - 40,6 км/ч*. С такой стремительностью дельфины могут двигаться лишь очень недолго (максимум несколько минут). Возможно, еще быстрее двигаются косатки, нападающие даже на быстроходных дельфинов. Японские ученые М. Нисиваки и Ч. Ханда в желудках этих хищников часто находили белокрылых морских свиней, высокие скоростные качества которых были установлены в опытных бассейнах Пойнт-Магу в Калифорнии и подтверждены данными физиологии: содержание кислорода в крови белокрылых морских свиней оказалось в 3 раза выше, а относительная масса сердца - в 2,4 раза больше, чем у афалины. По наблюдениям с судна "Монтерей", замерявшего быстроту хода некоторых видов китообразных, максимальная скорость плавания косаток в течение 20 мин варьировала от 38 до 55 км/ч. Последняя цифра, видимо, и лежит на грани гидродинамических возможностей отряда.
* (В 1966 г. Томас Ланг, оценивая гидродинамические свойства разных видов дельфинов, путем расчетов нашел ту скорость, при которой развивалась бы крайняя степень турбулентности - кавитация, совершенно исключающая дальнейшее повышение скорости при данной форме плавников. У обыкновенного дельфина это случилось бы при скорости 62, а у белокрылой морской свиньи - 74 км/ч)
Конечно, не у всех китообразных в одинаковой мере выражены скоростные качества. Наибольшая быстроходность свойственна рыбоядным формам, преследующим очень верткую и подвижную добычу (обыкновенные дельфины, продельфины, косатки, белокрылые морские свиньи, полосатики). Вместе с тем в отряде есть и настоящие тихоходы, питающиеся относительно малоподвижной пищей. Таковы бентосоядные серые киты, делающие лишь около 12 км/ч. А ловко прыгающие горбатые киты, планктоноядные гладкие киты и роющиеся на дне речные дельфины даже в рывках не превышают 18,5 км/ч. Но не они, а именно первая группа стоит в центре проблемы: какие приспособления обеспечивают ей высокую быстроходность в очень плотной среде.
Водная среда в 800 раз плотнее воздушной. Поэтому даже медленное передвижение в ней требует от водных обитателей обтекаемой формы тела. Вопрос о гидродинамической эффективности китообразных возник еще в 1936 г. в виде так называемого "парадокса Грэя". Английский ученый Джон Грэй, наблюдая за стремительным движением мелких китообразных в океане, высчитал, что из-за турбулентного потока мышцы дельфинов должны обладать мощностью, в семь раз большей, чем у всяких других млекопитающих. Поскольку это невозможно, то было высказано предположение, что дельфины имеют какие-то средства, помогающие им бороться с турбулентностью и преодолевать сопротивление воды. Особенно интенсивно этот вопрос изучался в последние годы.
Что помогло китообразным стать скороходами морей?
Конечно, это и обтекаемая форма тела с гладкой поверхностью, и совершенный локомоторный орган - сильный движитель, снабженный мощной мускулатурой с отличной энерговооруженностью и очень гибким хвостовым отделом позвоночника, и особая кожа, способная задерживать возникновение турбулентных пульсаций в пограничном слое воды, и, наконец, регулируемый гидроупругий эффект в плавниках. Познакомимся с этими адаптациями.
Форма тела китообразных торпедообразная, отлично обтекаемая, постепенно увеличивающаяся в толщине от кончика головы к грудному отделу и суживающаяся к хвосту. Тело заканчивается горизонтально расположенным хвостовым плавником в виде широкого равнобедренного треугольника, разделенного на заднем крае выемкой на две лопасти. Задние конечности исчезли, а передние превратились в жесткие весловидные грудные плавники, которые направляют животное вверх или вниз, а также помогают при поворотах и торможении. Имеющийся у многих видов спинной плавник, как стабилизатор, придает большую устойчивость телу в воде. Все плавники хорошо обтекаемы и в поперечном сечении имеют форму вытянутой падающей капли. Снаружи исчезло все, что мешает быстрому плаванию, в том числе волосяной покров (за исключением одиночных чувствующих волосков на голове), ушные раковины и мошонка. Под кожей образовался мощный теплозащитный слой жира. Исчезли сальные и потовые железы, а внешне незаметная пара млечных желез разместилась под кожей почти в эадней трети тела, по бокам (мочеполового отверстия. Сосок (у самцов отсутствует) скрыт в кожном кармане, откуда выступает лишь в лактационный период.
Голова, сидящая на короткой и жесткой шее, может наклоняться по отношению к туловищу до 45°, например у речных дельфинов, у которых все семь шейных позвонков свободные. У некоторых китообразных эти позвонки сливаются в единый шейный блок, и тогда голова становится неподвижной, как у гренландских китов. Тело всех представителей отряда исключительно гибкое в хвостовой части, где число позвонков увеличивается. Удлиненный хвостовой отдел позвоночника, в связи с редукцией крестцового отдела и таза, нечетко отграничен от поясничного отдела, и границы его на скелете могут быть установлены по углевидным (шевронным) косточкам, причленяющимся к хвостовым позвонкам снизу. Китообразные превосходно управляют своим главным локомоторным органом - хвостом. Хвостовой стебель совершает удары вверх-вниз, а лопасти работающего хвоста принимают разные углы наклона к продольной оси стебля: когда стебель идет вниз, лопасти поворачиваются вверх и наоборот. Поэтому хвостовой стебель сжат с боков. Вращательных движений при плавании хвост не производит. Частота и размах ударов хвоста и степень наклона лопастей влияют на скорость плавания. Во время плавания лопасти обычно не показываются, но их можно [видеть, когда животные целиком выпрыгивают из воды (все дельфины, полосатики). Некоторые же киты (гладкие, серые, кашалоты) специально выставляют из воды хвост, а гренландские им даже помахивают в воздухе. Горбачи во время акробатических, иногда 2-, 6-кратных прыжков размахивают своими очень длинными (до трети длины тела) грудными плавниками, а в воде способны сгибать конечность. В экспедиции Жака Кусто на судне "Калипсо" его сын Филипп снимал китов под водой и однажды проплыл с фотокамерой очень близко между самкой горбача и ее детенышем: тогда самка, чтобы не задеть аквалангиста, оттянула конец своего плавника.
Главный движитель китообразных - могучий хвост - приобрел необычайную силу в связи с коренным преобразованием их мускулатуры. Эти изменения подробно изучил В. А. Родионов. Он заметил отчетливое упрощение мышечной системы китообразных, насчитав у дельфинов лишь 112 мышц вместо 170 у наземных млекопитающих. Причины этого заключаются в глубоком адаптивном переустройстве отряда: в перемещении локомоторной функции на хвостовую часть тела, в редукции задних конечностей и в специализации передних (грудных) плавников, в слиянии одних мышц и исчезновении других, в изменении функций комплексов мышц соответственно требованиям водной среды.
В мышечной системе позвоночника особенно выделяется роль мощной двигательной мускулатуры, направленной на обеспечение быстрого передвижения китообразных в воде и достижение высокой маневренности. Упрощение мускулатуры позвоночника сопровождается гипертрофическим развитием двух пар мускульных комплексов (длиннейшей и гипаксимальной мышц), лежащих над и под позвоночным столбом и выполняющих основную нагрузку при работе хвостового движителя. Обе мышцы составляют почти 1/6 часть веса всех мышц тела. При этих условиях огромная масса мускулатуры позвоночного столба концентрирует свои усилия лишь на работе хвоста, на его ударах вверх-вниз (поэтому вес мышц, осуществляющих боковые изгибы хвоста, в 15 раз меньше веса мышц, двигающих хвостом в вертикальной плоскости). Эффективность работы мышц возрастает за счет очень высоких остистых отростков позвонков, к которым прикладывается усилие мышц при увеличенном плече рычага. Наконец, очень важно и то обстоятельство, что китообразные в воде находятся в состоянии, близком к невесомости; поэтому в отличие от наземных обитателей им не приходится тратить мускульную силу на преодоление собственного веса, и они расходуют ее целиком на движение как дополнительный источник энергии.
При макро- и микроскопическом изучении мускулатуры В. А. Родионов обратил внимание на соотношение красных и белых мышечных волокон и на распределение в них жира и гликогена. Красных волокон оказалось больше всего в самых активных мышцах, таких, как длиннейшая. Гликогена содержалось больше в красных, а жира в белых волокнах. Богатые миоглобином красные волокна действуют как своеобразный насос, который высасывает кислород из крови. При выключенном внешнем дыхании он экономно расходуется во время плавания. При исследовании иннервации мышц было обнаружено множество нервных окончаний и проприорецепторов в виде нервно-мышечных и нервно-сухожильных веретен. Столь богатая нервная аппаратура требуется для тонкой регуляции обменных процессов в мышце и для улучшенного восприятия положения тела в условиях ослабленной гравитации (сил тяготения) в воде.
Таким образом, В. А. Родионов, рассматривая секрет мощности хвостового движителя дельфинов, объяснил "парадокс Грэя" особенностями мускулатуры. Однако для этого одной идеальной формы тела и специфики мускулатуры оказалось явно недостаточно. Проблема превращения китообразных в лучших маневренных и скоростных пловцов была решена в ходе эволюции только тогда, когда у них появились еще два важнейших приспособления к быстрому плаванию: а) активная самонастройка демпфирования кожи и б) саморегуляция гидроупругости плавников.
Первая важнейшая адаптация китообразных заключается в рефлекторно регулируемом свойстве их кожи самонастраиваться - гасить турбулентные пульсации, возникающие в пограничном слое, и тем самым снижать гидродинамическое сопротивление. Это свойство демпфирования кожи и связанные с ним анатомические ее особенности были описаны советскими исследователями С. В. Першиным, В. Е. Соколовым, В. В. Бабенко, Л. Ф. Козловым и другими. Демпфирование кожи в основном осуществляется сосочковым слоем, обильно снабженным кровеносными сосудами и нервами. Каждый сосочек кожи благодаря увеличению или сужению просвета кровеносных сосудов на различных скоростях плавания обладает переменной упругостью. Выполнению указанной функции соответствуют и направленные вдоль струй потока дермальные валики, которые служат основанием мельчайших сосочков кожи.
О ламинаризации потока кожей китообразных свидетельствуют и некоторые прямые наблюдения над дельфинами, плавающими в условиях биолюминесценции (свечения) моря. Еще в годы второй мировой войны английский биолог Г. А. Стефен, служивший на флоте, наблюдал замечательный факт: ночью при тихой погоде в светящемся море за стремительно плывущим дельфином остаются лишь два огненных шнура, за плывущим тюленем - широкое полыхающее поле. Аналогичные наблюдения были проведены в 1967 г. в водах Калифорнии с исследовательского судна-катамарана "Си-си" ("Смотри море"), принадлежащего американскому центру подводных исследований: из прозрачного трехметрового шара, опускающегося под днище катамарана, Вильям Эванс и Лари Мак-Кинли ночью отлично видели, как у быстро плывущих короткоголовых дельфинов от кончиков спинного и обоих грудных плавников протягивалась назад светящаяся узкая полоска и три такие же узкие полоски тянулись от двух уголков и серединной выемки хвостового плавника. Следовательно, дельфин плывет, почти не нарушая спокойного состояния воды.
Высокие гидродинамические свойства кожи дельфинов ученые пытаются использовать в технике для моделирования и создания обшивок скоростных кораблей. К таким попыткам следует отнести, например, создание немецким гидродинамиком Максом Крамером обшивки "ламинфло". Другие ищут средства повышения скорости кораблей, повторяя форму китообразных в контурах, например, подводных лодок. Третьи пытаются снизить гидродинамическое сопротивление, оказывая влияние на плывущее тело различными растворителями, полимерными реологическими жидкостями, подражая в какой-то мере слизи рыб. Над проблемой применения полимеров в гидродинамических целях в настоящее время работают исследователи ряда стран (США, Англии, ФРГ).
Вторая важнейшая адаптация к быстрому плаванию китообразных - переменная гидроупругость плавников, регулируемая в зависимости от режима плавания. Она была установлена советскими учеными С. В. Першиным, А. С. Соколовым и А. Г. Томилиным в 1968 г. на основе разносторонних (морфофункциональных, гидродинамических, гемодинамических и экологических) исследований пяти видов дельфинов (афалины, белобочки, азовки, белухи, белокрылой морской свиньи) и трех видов китов (кашалота, финвала и сейвала). Ученые выполнили гидродинамический анализ подводных киносъемок плавания дельфинов в естественных условиях и в разных ситуациях, проанализировали рентгенограммы сосудистой сети и гистологических препаратов тканей плавников дельфинов и китов, провели натурные эксперименты и техническое моделирование явления. Регуляция упругости плавников у китообразных осуществляется автоматически при помощи специфических комплексных артериовенозных сосудов, распределительных узлов на их системе и особой структуры покровных тканей плавников, включающей покрытие из сухожильных тяжей. Чем быстрее дельфины плывут, тем выше упругость их плавников. Прямое доказательство этого явления можно видеть на косатках в океанариях: когда касатки малоподвижны, их весьма высокий спинной плавник сгибается на 180°, свисая вершиной прямо вниз; но стоит им быстро поплыть или совершить энергичные прыжки, как спинной плавник обретает упругость и выпрямляется.
Хвостовой плавник - широкий, сильно развитый вырост кожи - сохраняет ее типичную трехслойную структуру : эпидермиса, дермы и гиподермы. Но есть в этом выросте различия по сравнению с кожей туловища:
а) между сосочковым слоем дермы и сетчатым слоем гиподермы залегает сплошное и очень малорастяжимое покрытие. Оно состоит из многочисленных сухожильных тяжей (в диаметре 1-1,5 мм), которые тянутся от туловищных мышц и хвостовых позвонков;
б) сетчатая гиподерма с коллагеновыми волокнами, заполняющая всю внутреннюю область плавника, весьма бедна жировыми клетками, но очень насыщена кровеносными сосудами.
Хвостовой плавник (рис. 15, А) не имеет собственной мускулатуры, но его кровоснабжение очень обильно: мощная сеть артерий и вен всех рангов, от крупных распределителей и собирателей (в диаметре 1-2 мм) до капилляров (0,02-0,04 мм). При работе сердца артериальные сосуды могут сильно изменять объем сосудистого русла, свою емкость и пропускную способность. При большой физической нагрузке минутный расход крови может повышаться десятикратно за счет кровяных депо и увеличения калибра сосудов. В сети капилляров с изменением кровесодержания развивается осмотическое давление до нескольких атмосфер.
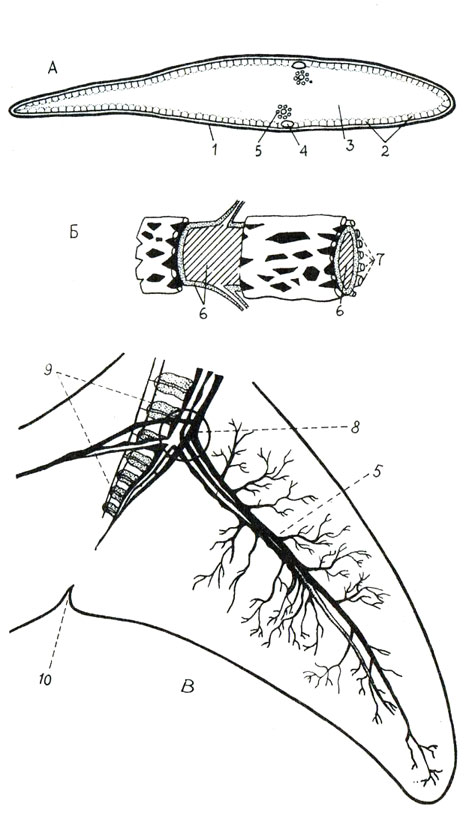
Рис. 15. Хвостовой плавник дельфинов - это и локомоторный орган с переменной упругостью и теплорегулятор: А - поперечный разрез через хвостовую лопасть дельфина: 1 - кожа, 2 - слой сухожильных тяжей, 3 - гиподерма с коллагеновыми волокнами, 4 - одиночная вена, 5 - магистральный комплексный сосуд; - комплексный сосуд; 6 - центральная толстостенная артерия, 7 - тонкостенные вены, окружающие артерию; В - хвостовая лопасть с магистральным типом ветвления сосудистой сети: 5 - магистральный комплексный сосуд, 8 - распределительный узел потока крови, 9 - последние хвостовые позвонки, 10 - выемка хвостового плавника
Специфическая особенность сосудистой сети плавников - ее комплексный характер. Комплексный сосуд состоит из толстостенной мышечной артерии и венозной оплетки - тонкостенных вен, которые окружают артерию (рис. 15, Б). Венозная оплетка не сплошь покрывает поверхность артерии, а со значительными просветами. Число вен в оплетке достигает десяти (редко больше). Обилие анастамозов в венозной оплетке связано с регулированием давления и интенсивности кровотока: повышение или понижение давления в артерии ведет к расширению или сокращению ее стенки, а это изменяет давление на венозную оплетку и режим ее работы, вовлекая в действие и капиллярную сеть в гиподерме.
Таким образом, кровь, поступая по комплексным сосудам в плавники то в большем количестве (при высокой скорости животного), то в меньшем (при малой скорости), создает внутри плавников, одетых в нерастяжимую оболочку, различную степень упругости, объемное напряженное состояние, связанное с режимом плавания.
Мощно развитые мускулистые стенки в артерии комплексного сосуда позволяют тонко регулировать количество поступающей крови, а присутствие одиночных вен с утолщенными стенками указывает на возможность дополнительного ее оттока в тех случаях, когда она нагнетается более энергично.
Главные артерии лопастей хвостового плавника отделяются от одного магистрального хвостового сосуда между позвонками (рис. 15, В). В местах их разветвления на магистральной артерии образуются распределительные узлы, от каждого из которых отходят по четыре боковых сосуда: по два в правую и левую лопасти, сверху и снизу. При этом могут быть три типа ветвления :магистральный (у мелких и быстро плавающих дельфинов), когда резко выделяется один распределительный узел; отходящие от этого узла сосуды питают почти всю лопасть; рассыпной - у крупных китов-тихоходов (кашалотов) и тихоходов-дельфинов ((белухи); у них есть серии последовательно расположенных многочисленных узлов; полурассыпной (промежуточный) - у крупных китов-быстроходов (полосатиков - финвала и сейвала); у них среди 7-12 ветвей выделяются 2-3 наиболее крупных узла.
При магистральном типе ветвления регуляция скорости и давления крови в плавниках более совершенная, чем при рассыпном. Когда хвостовые лопасти достигают огромных размеров, сосуды не могут увеличиться пропорционально росту лопастей, так как при их большом калибре создаются условия для турбулизации потока крови, и в этом случае рассыпной тип оказывается совершеннее магистрального. Таким образом, типы ветвления главных сосудов в хвостовых лопастях определяются как размерами животных, так и их скоростными качествами. В разных частях хвостовой лопасти опорные элементы тканей - коллагеновые и эластиновые волокна расположены по-разному: в передней толстой части профиля лопастей волокна направлены беспорядочно, а в тонкой (концевой) части - вертикально. Это, очевидно, связано с необходимостью иметь разную эластичность в разных частях плавника. Регулируемый гидроупругий эффект в плавниках помогает китообразным двигаться с большой скоростью, преодолевая огромные расстояния, и проводить всю жизнь в непрерывном движении - ныряниях (для лова добычи) и подъемах к поверхности (для дыхания).
Сосудистая система в плавниках китообразных выполняет еще одну важную функцию - осуществляет регуляцию тепла.
|
ПОИСК:
|
При использовании материалов проекта активная ссылка обязательна:
http://underwater.su/ 'Человек и подводный мир'